
涉刑当事人及家属必备手册(八)——检察院阶段争取不起诉或缓刑
涉刑当事人及家属必备手册(八)——检察院阶段争取不起诉或缓刑 在前两期中,我们为大家介绍了检察院审查起诉阶段的大致流程,辩护人需要做的包括阅卷、核实、收集、申请调取...
2026-01-09 18:19:30

合弓纲与蜥形纲的竞争史(下篇)
二叠纪——三叠纪大灭绝事件是地质史上最严重的大灭绝事件,当时地球上70%的陆生脊椎动物,以及高达96%的海中生物消失。而这一次大灭绝事件,也可以说从根本上逆转了合弓纲与蜥形纲的竞争状况。
由于三叠纪以史上最大的一次灭绝事件开始,空出了大量生态位,因此从一开始起其生物分化就很厉害(三叠纪以地层被明显地分为上下三段而得名)。在二叠纪完全没有存在感的主龙类,一到了三叠纪就迅速展开了成功的辐射进化。
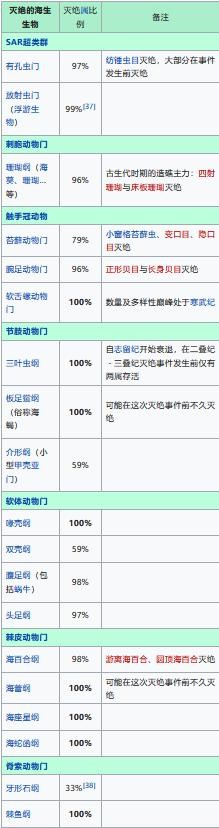
二叠纪——三叠纪大灭绝事件的物种灭绝情况

在三叠纪初的某些地层,水龙兽占了动物化石的90%。试想一觉醒来遍地都是这种像猪一样的食草动物的情形。
在三叠纪,主龙类快速地成为陆地上的优势脊椎动物。关于主龙类为何快速崛起,最常提出的两个解释分别为:
1、主龙类演化出直立四肢的过程,比合弓类的演化还快;这让它们有更强的耐力,可以在运动时同时呼吸。但这论点是有问题的,因为主龙类拥有优势地位时,它们仍处于躺卧或半直立姿势,类似水龙兽或其他合弓类。
2、三叠纪早期的气候环境普遍干燥,这是因为当时的陆地集合为盘古大陆,而主龙类的保存水分能力比合弓类好。现代双孔动物(蜥蜴、蛇、鳄鱼、鸟类)都将尿酸以糊状排泄出来,根据合理的推测,主龙类也将尿酸排泄出来,因此能更好地保存水分。而双孔动物缺乏皮腺的皮肤也有助于保存水分。现代哺乳类排泄尿素,需要大量的水分以溶解尿素,皮肤也布有许多皮腺,会流失许多水分,这就难以适应三叠纪时期极其干燥的气候,因此它们在干燥的环境中呈现出劣势。在澳洲的一个地层显示出澳洲的上新世至更新世期间,气候可能一样干燥,而该地最大型的陆地掠食动物是古巨蜥,而非哺乳类。
不过,蜥形纲对合弓纲霸主地位的取代也不是瞬间就完成的,至少在三叠纪的前期,合弓纲仍然有一定的繁盛度。二齿兽下目的水龙兽在二叠纪-三叠纪灭绝事件中存活下来的原因,可能是因为它们的祖先体型娇小、动作灵活而且居住在地下,比其他大型动物更快适应灭绝事件后的环境。而从水龙兽进化而来的肯氏兽,体型上远大于水龙兽,其结实肩带、骨盘可协助支撑2—4米长的身体,拥有强力的嘴部还有强壮的下颌肌肉,可切碎植物,是适应良好的草食性动物,类似于今天的水牛。虽然它们的头部很大,但因为眼窝与鼻腔的尺寸关系,重量却很轻运动却很灵活。肯氏兽很快取代了水龙兽的生态位。到了三叠纪中期,肯氏兽是广布全世界的动物。
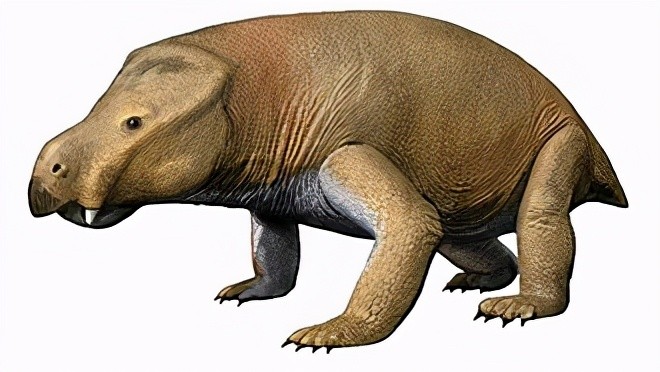
宽头肯氏兽

集群的肯氏兽
犬齿兽亚目是兽孔目中最多样性的其中一群。它们以类似狗的牙齿命名。而犬齿兽类在三叠纪也经历了进一步的演化,其中真犬齿兽下目的牙齿经过特化,使它们有不同的作用,包括撕裂与咀嚼,犬颌兽为其中最原始的类型。到了奇尼瓜齿兽总科这个子类,体型接近于狗,已经具有许多类似哺乳动物的特征,有六个后犬齿,其后齿是多尖头的,而主要的尖齿向后弯曲,具有高度弓形的颧弓,最早出现于晚三叠世的卡尼阶。而今天的哺乳纲也属于广义上的犬齿兽类,从分法上来看可以归纳在“合弓纲—兽孔目—真兽孔类—新兽孔类—兽齿类—真兽齿类—犬齿兽亚目—真犬齿兽下目—新颌兽小目—奇尼瓜齿兽总科—哺乳形类—哺乳纲”下。在约2.3亿年前的三叠纪晚期,早期的哺乳动物与恐龙几乎在同一时期正式出现了。
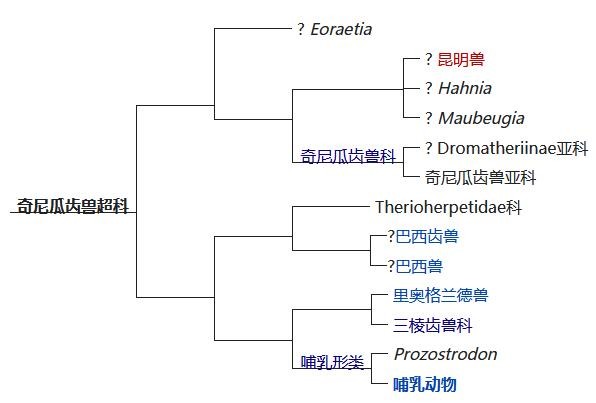
奇尼瓜齿兽总科下的演化分支
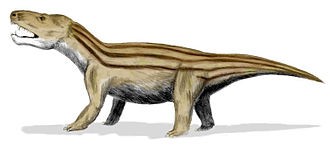
犬颌兽是三叠纪中期最类似哺乳类的真犬齿兽类

哺乳形类的Probelesodon

奇尼瓜齿兽
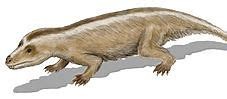
与哺乳动物关系极近的三棱齿兽科,生活于晚三叠纪到中侏罗纪,为小型肉食性或食虫性真犬齿兽类。

兽头亚目虽有少数躲过二叠纪末的大灭绝,但在三叠纪中期就渐趋绝迹,图为兽头类化石中年代最晚的幻嵌齿兽。

晚三叠纪开始出现的华氏摩尔根兽复原图

三叠纪晚期的原始哺乳类冈瓦那兽

三叠纪晚期的另一类原始哺乳类贼兽

真贼兽类
槽齿目是个原始主龙类的并系群集合,也被视为恐龙、翼龙、鳄鱼的共同祖先和三叠纪时期的霸主,但这个分类目前已过时而被淘汰。槽齿目的定义是特定的共同原始特征,例如:眶前孔(一对在头骨两旁介于眼窝与鼻孔间的洞)与位在槽洞里的牙齿。槽齿指的是槽齿目的牙齿都位在颌骨的槽洞中;这是一个主龙类的特征,而恐龙仍保有这个特征。由于这个并系群内的亲缘关系较为复杂,目前已不再简单地视为一类。槽齿类中从二叠纪末开始出现的最原始的古鳄科再到三叠纪晚期的劳氏鳄、波波龙等只能根据具体的亲缘关系划分在主龙形类和主龙类演化支系的不同位置。槽齿类中与鳄鱼关系较近的部分直接划为伪鳄类,伪鳄类是三叠纪晚期最主要的统治者。

主龙形类的演化分支
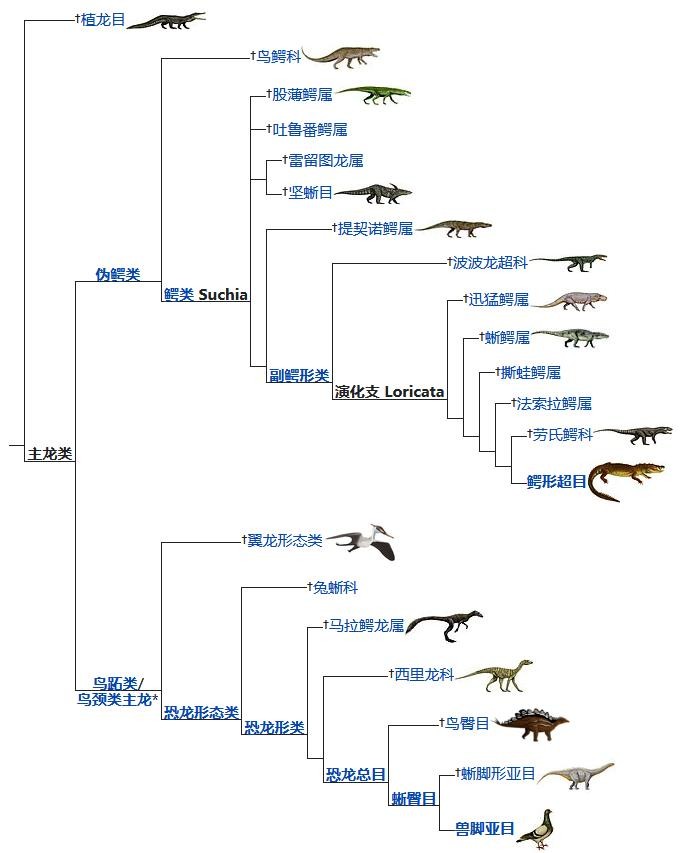
主龙类的演化分支
在三叠纪中早期,由古鳄科辐射进化而来的引鳄、黄昏鳄、加斯马吐鳄、派克鳄、滨鳄、迦梨鳄、原鳄龙、武氏龙、吐鲁番鳄等迅速繁盛于陆生水域地区。其中引鳄科和武氏龙属中较大的已能达到五六米长,是当时的大型掠食动物。
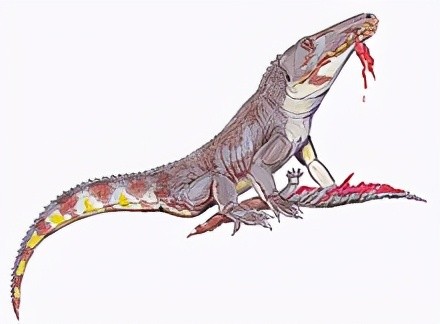
加斯马吐鳄的上颌前端往下弯曲,可协助咬住猎物,腭骨则有一排牙齿。后期主龙类失去这两个原始特征。

匍匐着的引鳄
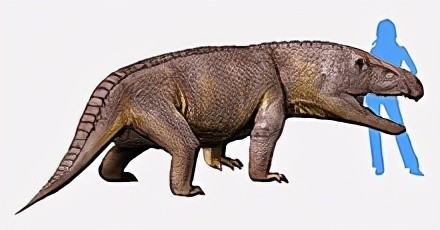
引鳄复原图
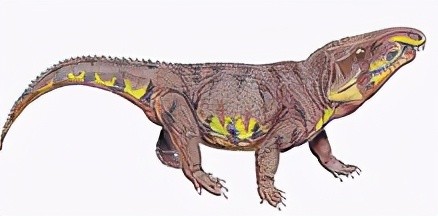
生活于三叠纪中期的俄罗斯的一种引鳄Chalishevia
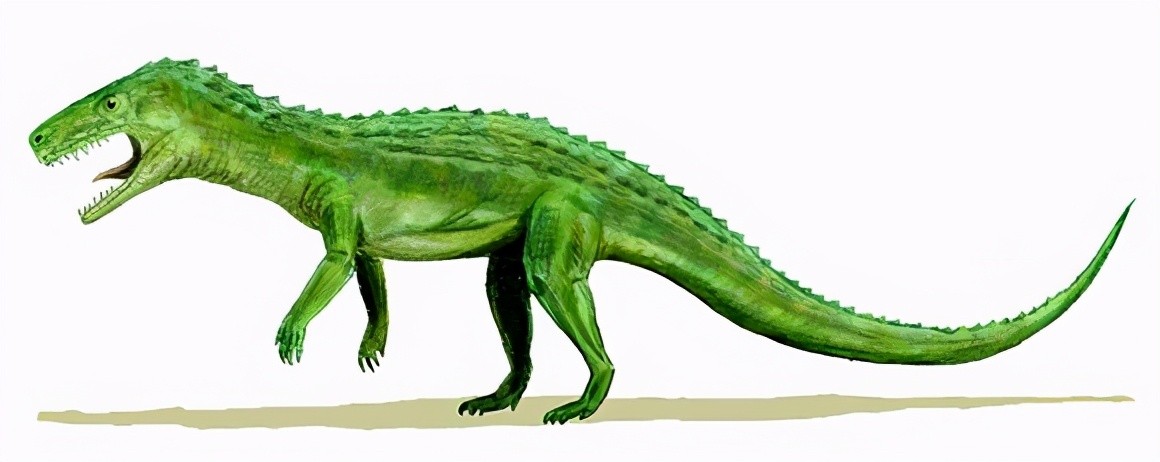
派克鳄
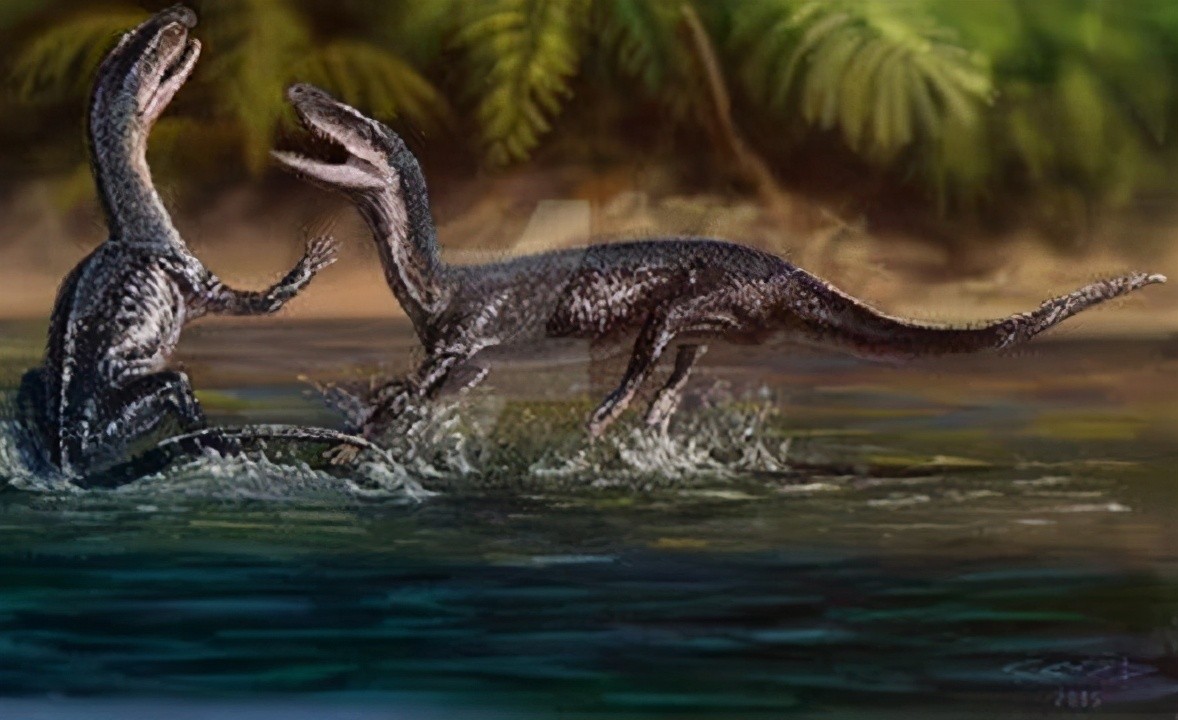
生活于三叠纪中期的戏水的吐鲁番鳄
伪鳄类是主龙形下纲中的两个主要演化支之一,它们的头骨通常厚重,尤其是与鸟颈类主龙相比;伪鳄类主龙的口鼻部通常是长而狭窄的,颈部短而强壮,四肢的结构介于典型爬行动物的往两侧延伸到恐龙或哺乳类的直立四肢之间(虽然恐龙与哺乳类的姿态不同)。身体通常由两排或更多排的甲板保护者。伪鳄类主龙的体形大多数相当大,通常可达3米以上,最大的甚至已达到了十几米长,遍布三叠纪的水域和陆地。

波斯特鳄是现代鳄鱼的早期远亲,长约4—6米,可能猎食其他较小型的爬行动物。

法索拉鳄长达8—10米,生活于三叠纪晚期的诺利阶。

蜥鳄体长约5—7米,是除法索拉鳄以外的最大型副鳄形类,以完全直立的四肢行走并可能以埋伏方式攻击猎物。

波波龙的亲缘关系接近现代鳄鱼,离恐龙的关系较远。它可能是在主龙类的早期演化阶段独自演化出了二足步态。
蜥形纲在三叠纪的崛起也同样繁盛于海洋地区。目前还不是很明确称霸于海洋的鳍龙总目是由怎样的演化路线而来。而现代龟鳖已经在遗传分子学研究中被认为属于主龙形类,其与鳍龙总目的楯齿龙目在身体结构和形态上又较为接近,楯齿龙类身长通常为1到2米,最大型的物种可达3米。它们拥有短而非常强壮的四肢。部分原始物种在外表上类似粗厚、水桶腰的蜥蜴,而其他的物种则因为背上的大型骨板,而类似乌龟,估计龟鳖目可能是在二叠纪末至三叠纪初从楯齿龙类的较近祖先分化而来。
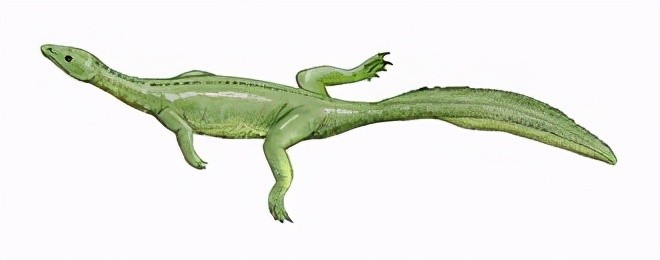
贫齿龙牙齿稀少且局限于上下颌前端,并非纯肉食性,所属的海龙目形如蜥蜴,演化位置介于鱼龙与主龙形下纲。
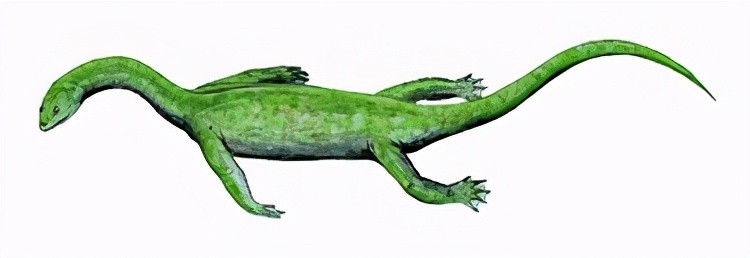
幻龙目中较原始的肿肋龙亚目,颌部前部布满钉状牙齿,其以捕食海洋鱼类为生,体长20厘米至一米多。

幻龙科头部长、宽广、平坦,用排列着针状牙齿的长颌捕抓鱼类与其他海中动物,可能先缓慢跟踪猎物然后突袭。

生活于三叠纪晚期的楯齿龙目的楯齿属,在外形上类似于海生龟鳖。

2.51亿多年前的三叠纪初的龟山巢湖龙是现已知最早的鱼龙类,身长仅70到170公分,重约10公斤。

皮氏吐龙长约3米,同时拥有幻龙类的腭骨与身体形状与蛇颈龙类僵直的脊椎骨的特征,为两者之间的过渡物种。
而三叠纪时期合弓纲的衰落和蜥形纲的崛起是一个逐渐的过程,在晚三叠世的卡尼期的两次小型灭绝事件——兰格利亚火山爆发事件和卡尼期洪积事件同样起到了很重要的作用。约2.35亿年前大规模的兰格利亚火山喷发将二氧化碳、甲烷和水蒸气排入大气,导致全球变暖和全球降雨增加。而2.34亿到2.32亿年前的卡尼期洪积事件导致地球明显变得更加潮湿,于牙形磷灰石中发现的氧稳定同位素(δ18O)的约1.5‰负移表明该时期出现了全球性的升温,负责生产碳酸钙的生物发生了重大变化,菊石、牙形石、苔藓虫和海百合的灭绝率在此时期增高。这两个事件在客观上都导致了鸟颈类主龙(恐龙、翼龙)和哺乳形类的崛起,恐龙类刚开始出现时仅占脊椎动物化石中的不足2%,但在这两次事件之后迅速发展并逐步取代部分伪鳄类的生态位,而哺乳形类在与新兴更先进的蜥形纲类型的竞争中进一步地落于下风。
三叠纪-侏罗纪灭绝事件则作为又一场灭绝事件进一步拉大了合弓纲与蜥形纲之间的落差,本次大灭绝中牙形石灭绝,除鱼龙和蛇颈龙外所有的海生爬行动物消失。腕足动物、腹足动物和贝壳等无脊椎动物受到巨大冲击。在海洋中,22%的属,大约一半的种消失。大多数合弓类动物伴随着以伪鳄类和鳍龙类为主的非鸟颈类主龙的主龙形类一同消失了。而在侏罗纪和白垩纪,合弓纲的境遇自然比二叠纪的蜥形纲还要更为不堪。这些早期的原始有胎盘类,大多体型缩小到只有老鼠般大而可能仅以昆虫为食,而且通常夜晚行动以尽可能避免天敌的威胁。
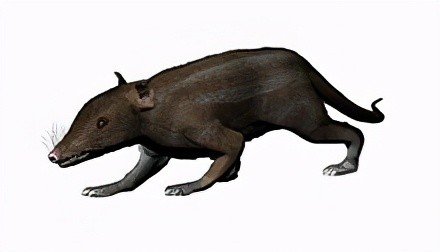
中华侏罗兽化石只留下了约2.2厘米的不完整头骨、部分头后骨架,及毛发等软组织的印痕,可能以昆虫为食。

中国袋兽是后兽下纲中最古老的化石,在白垩纪初, 只有15厘米长,可能重30克,可能以昆虫为食。

獭形狸尾兽为已知最大的侏罗纪和白垩纪的兽类,可能重约500-800克及最少长42.5厘米,捕鱼为生。
白垩纪——第三纪大灭绝事件之后,历史则逐渐再一次返回到了古生代晚期的状态,合弓纲又重新兴起。恐龙后裔的大型食肉鸟类在新生代也曾零星出现过一些较有战斗力的种类,如恐鹤科中的泰坦鸟,也曾在数百万年前称霸于美洲大陆,能够捕食各种中小型哺乳动物,再比如灭绝于18世纪的新西兰的恐鸟和和17世纪以前存在于马达加斯加的象鸟。

泰坦鸟捕食

恐鸟

象鸟
现存哺乳类的大脑机制与鸟类仍有明显的不同,哺乳类在进化过程中演化出了发达的大脑皮层,成为思维能力的主要来源。而鸟类的大脑皮质不是很发达,更多地依赖大脑基底神经节的纹状体,纹状体作为鸟类的控制中心操纵着鸟类的各种活动。鸟类的大脑一般都非常小,比如渡鸦的大脑只有核桃般大,但是其中的纹状体却非常发达,在同等体积质量下的效率却要远高于哺乳动物的大脑皮层。鸟类的大脑神经密度是哺乳动物的3~4倍,鸦科大脑重量只有黑猩猩的二十六分之一,但却能通过镜子测试而与黑猩猩在智力上不相上下。鸟类的智力主要就取决于纹状体的发达程度,鸦科和鹦鹉科虽然脑容量不及鸡和鸠鸽科但却比后两者要聪明。鸟类的记忆效率则高得惊人,例如克拉克星鸦每年会将22000~33000颗谷粒埋在约15平方英里面积内,而观察发现它们时隔一年还能找到约5000处埋藏地点,超越了人类(实验证明时隔一个月人类就只能记住约2500处埋藏地点了)。从这个意义上讲,蜥形纲(龙族)的最先进物种相对于合弓纲(兽族)的最先进物种仍然存在一定方面的优势。
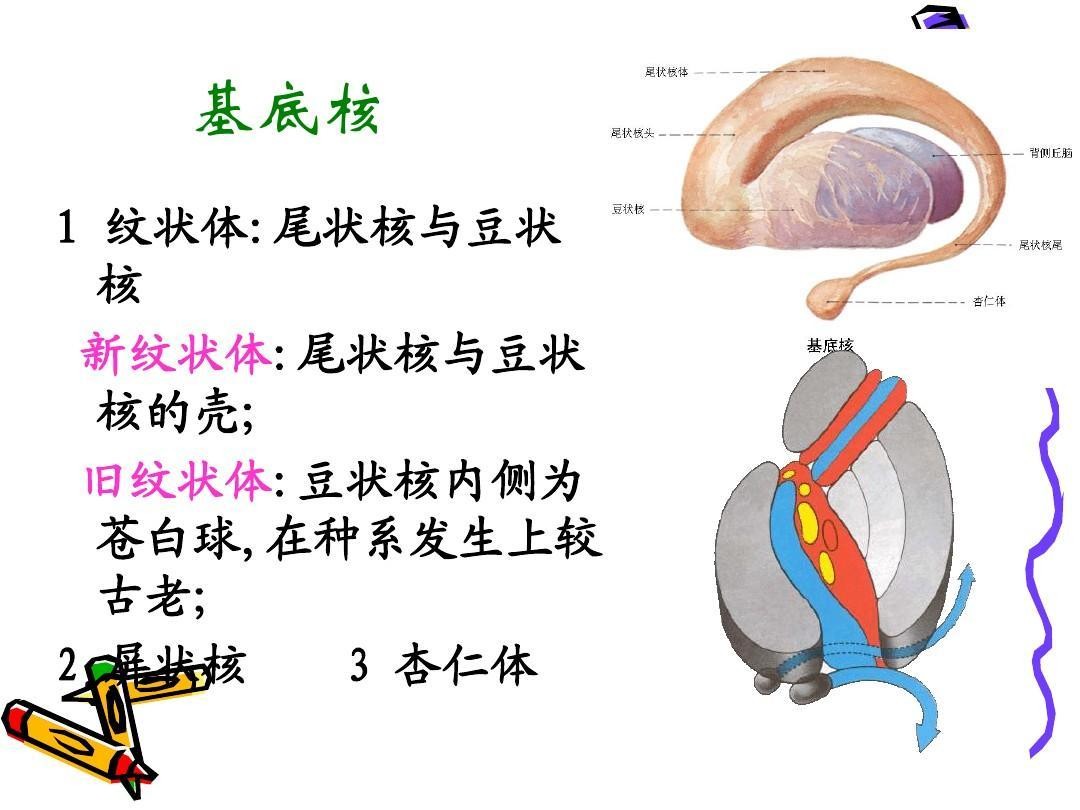
纹状体

灵长类以脑桥核发挥智能,鸟类的则很小,用类似结构的内侧螺旋核在皮质和小脑间传递信息来计划复杂的行为。
但是鸟类之于恐龙类而言本身就已是其中高度特化的类型,而鸟类中躲过这次大灭绝的也只有少数几个会筑巢的种类,在巢穴中躲过了天灾,这也导致了当代鸟类的基因多样性非常之差,特化程度过高而难以再次大范围辐射进化。鸟类虽然有一万多种,哺乳动物只有几千种,但鸟类的基因多样性程度还比不上哺乳纲的一个食肉目。但总的来说鸟类虽然是蜥形纲中最进化的类群,但一类物种能在以后崛起的首要前提在于自身的基因多样性而带来的潜力,特化程度低才能突变方向多,从这个意义上讲,蜥形纲的未来其实还是在种类本身就丰富多样的蜥蜴身上。
2026-01-09 18:19:30

2026-01-09 18:17:16

2026-01-09 18:15:02

2026-01-09 18:12:47

2026-01-09 18:10:33

2026-01-09 18:08:19

2026-01-09 18:06:04

2026-01-09 18:03:50

2026-01-09 18:01:36

2026-01-09 17:59:21

2026-01-09 17:57:07

2026-01-09 17:54:53

2026-01-09 03:00:06

2026-01-09 02:57:51

2026-01-09 02:53:23

2026-01-09 02:51:08

2026-01-09 02:48:54

2026-01-09 02:46:40

2026-01-09 02:44:25